7月9日,Nature Communications在线发表了华中农业大学棉花遗传改良团队题为“Evolution and subfunctionalization ofCIPK6homologous genes in regulating cotton drought resistance”的研究论文。该研究揭示了异源四倍体陆地棉中CIPK6同源基因出现功能分化,形成GhCBL1A1-GhCIPK6D1和GhCBL2A1-GhCIPK6D3两个信号通路分别正、负调控棉花抗旱性的分子机制,为多倍体植物中同源基因调控生长发育和响应逆境的亚功能化提供了新视角。

棉花是重要的经济作物和油料作物。随着我国作物种植布局的变化,棉花种植区域逐渐向高温、干旱地区转移,在新一轮农业产业结构调整中,新疆棉区成为我国最重要的棉区。棉花生育期长,生长盛期又值高温季节,蒸腾作用强,需水量大,干旱胁迫成为影响棉花生产的主要制约因素之一。发掘抗旱基因、解析抗旱机制并创造抗旱种质是棉花抗旱育种的重要目标。
棉花团队前期在陆地棉中系统鉴定了CBLs和CIPKs基因家族成员(Sun et al. 2021)。从物种进化的角度出发,结合亚洲棉和雷蒙德氏棉两个二倍体棉属,对CIPK6s基因家族的8个成员进行了从头进化分析,推演了8个成员进化的顺序,发现CIPK6A1/D1是最古老的成员,而CIPK6A3/D3是基因组复制后产生的新成员。对亚洲棉、雷蒙德氏棉和陆地棉中的CIPK6s基因表达模式分析发现,四倍化后,GhCIPK6s基因受干旱诱导的表达模式呈现差异性。作者发掘到两个受干旱诱导显著上调表达的基因GhCIPK6D1和GhCIPK6D3。其中GhCIPK6D1本底表达极低,受干旱胁迫后急剧上调表达;而GhCIPK6D3是8个成员中本底表达量最高的,受干旱诱导后也显著上调表达。
作者创制了GhCIPK6D1和GhCIPK6D3稳定遗传的超表达和干涉/敲除转基因株系开展研究。发现超表达GhCIPK6D1株系敏旱,突变GhCIPK6D1株系抗旱;而GhCIPPK6D3表现相反,超表达株系抗旱,干涉株系敏旱。这表明在陆地棉中GhCIPK6D1和GhCIPK6D3在调控棉花抗旱性上出现了功能分化,GhCIPK6D1负调控棉花抗旱性,GhCIPK6D3正调控棉花抗旱性(图1)。
进一步研究发现转基因植株抗旱性和水分的散失有关,通过扫描电镜对转基因材料在干旱前后叶片气孔观察发现,正常情况下,不同材料间气孔开度没有显著差异;干旱胁迫下,与对照植株相比,超表达GhCIPK6D1株系气孔开度显著增大,突变GhCIPK6D1株系气孔开度显著减小,表明GhCIPK6D1负调控干旱胁迫下棉花叶片气孔开度。而GhCIPK6D3呈现相反表型,正调控干旱胁迫下棉花叶片气孔开度。进一步通过非损伤微测技术(NMT)对保卫细胞钾离子流进行测定,发现GhCIPK6D1抑制K+ 外排,使植株敏旱;GhCIPK6D3促进K+ 外排,使植株抗旱(图1)。
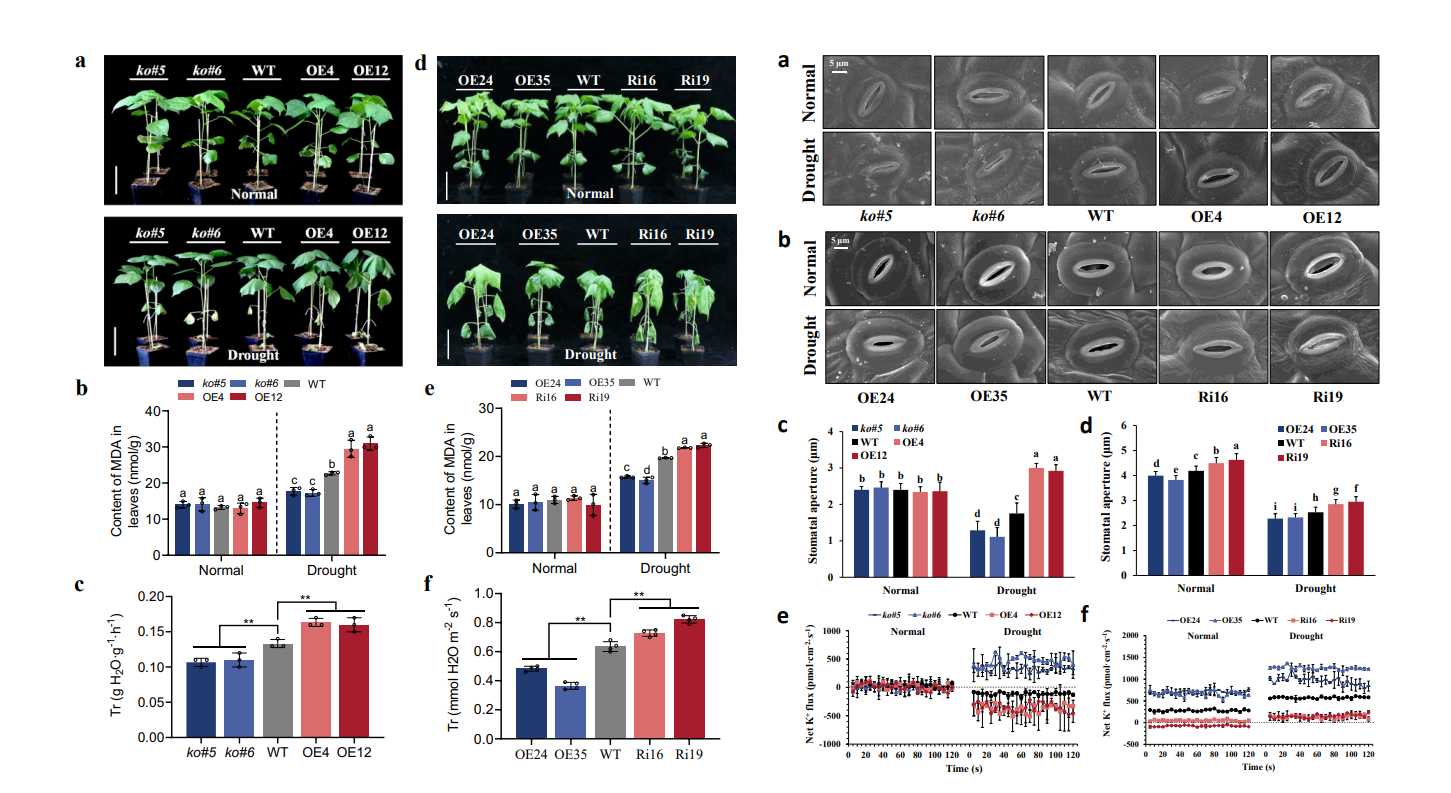
图1 GhCIPK6D1负调控和GhCIPK6D3正调控棉花抗旱性
CBL-CIPK信号系统是响应逆境胁迫的核心模块。作者对CBL1/2s和CIPK6s蛋白成员在亚洲棉、雷蒙德氏棉和陆地棉中互作模式进行分析,发现四倍化后,CBL1/2-CIPK6的互作模式在二倍体和四倍体中出现较大变化:雷蒙德氏棉中,GrCIPK6D1和GrCIPK6D3与GrCBL1/2均不发生互作,而多倍化后,出现GhCIPK6D1与GhCBL1A1互作;GhCIPK6D3与GhCBL1/CBL2的不同成员出现互作。结合团队前期的工作(Deng et al. 2020, Plant Physiology),通过多种手段验证了GhCBL1A1-GhCIPK6D1和GhCBL2-GhCIPK6D3体内体外的互作,证明了GhCIPK6D1被GhCBL1A1招募定位于质膜,而GhCIPK6D3被GhCBL2A1招募定位于液泡膜。
研究创制了GhCBL1A1和GhCBL2A1稳定遗传的超表达和干涉/敲除转基因株系解析CBL1A1-GhCIPK6D1和GhCBL2A1-GhCIPK6D3调控棉花响应干旱的生物学功能,发现GhCBL1A1通过负调控气孔开度负调控棉花抗旱性,GhCBL2A1通过正调控气孔开度正调控棉花抗旱性,与GhCIPK6D1和GhCIPK6D3功能协同。作者进一步利用遗传互补材料,验证了GhCBL1A1-GhCIPK6D1协同负调控气孔开度负调控棉花抗旱性,及GhCBL2A1-GhCIPK6D3协同正调控气孔开度正调控棉花抗旱性,且GhCIPK6D3的调控抗旱的生物学功能不依赖于GhCBL1。
该研究揭示了陆地棉CIPK6s基因成员中由于多倍化及基因组复制形成的同源基因调控棉花抗旱性的功能分化现象,并解析了其调控机制。GhCBL1A1-GhCIPK6D1和GhCBL2A1-GhCIPK6D3两个模块都可以通过调控气孔运动来调控棉花抗旱性。然而两者调控机制不同,GhCIPK6D1被质膜定位的GhCBL1A1招募,磷酸化质膜定位的钾通道,抑制K+ 在质膜上的外排,进而调控气孔运动,从而负调控棉花抗旱性;而GhCIPK6D3被液泡膜定位的GhCBL2A1招募,磷酸化液泡膜定位的钾离子通道蛋白,促进胞质中的K+ 离子进入液泡,进而调控气孔运动,从而正调控棉花抗旱性(图2)。
该研究结果不仅为CBL-CIPK信号系统的相互作用及参与气孔运动在干旱胁迫下的进化而调控抗旱性提供了不同的见解。对对棉花抗旱的遗传改良及抗旱育种有着重要的理论意义。
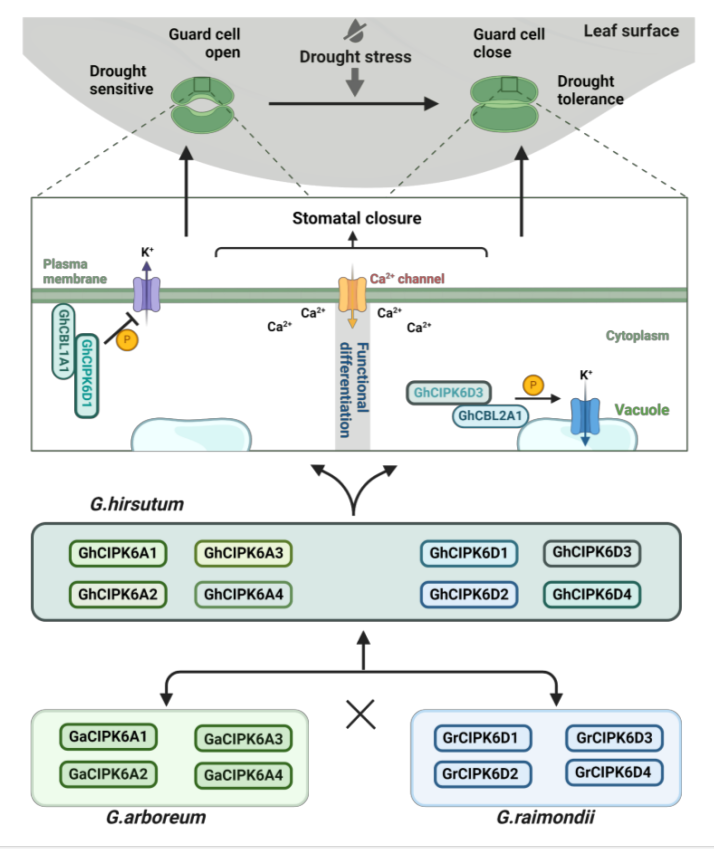
图2 GhCIPK6D1和GhCIPK6D3调控棉花抗旱性功能分化模式图
华中农业大学作物遗传改良全国重点实验室棉花遗传改良团队、湖北洪山实验室博士后孙伟男和博士生夏林杰为论文共同第一作者,棉花团队杨细燕教授为论文通讯作者,团队学术带头人张献龙院士参与了该研究的设计和指导,团队朱龙付教授、金双侠教授、王茂军教授和英国杜伦大学Keith Lindsey教授也参与了该项工作。该研究得到了国家自然科学基金面上项目、国家自然科学基金青年基金、国家重点研发计划和新疆兵团科技项目的资助。
