近日,我校水稻科研团队在研究中解析了水稻适应长日照成花的分子复合物模型,为调控水稻品种地域适应性、提高产量提供了新的途径。
植物开花调控的分子机制比较保守,一般在叶片的韧皮部形成“开花素”(florigen)。“开花素”经过维管束运输到茎顶端分生组织形成成花素激活复合物(Florigen activation complex, FAC),启动植物由营养生长转向生殖生长。因此,成花素激活复合物的形成决定了植物开花的时间及农作物品种种植的区域。水稻作为重要的粮食作物和基因组研究的模式植物,抽穗期决定了品种的季节和地区适应性,是影响其产量的重要因素之一。
虽然水稻在短日照条件下容易诱导其抽穗(成花转换),但经历长期的人工选择,培育出了大量适宜在高纬度(长日照条件)适时抽穗的水稻品种。水稻基因组中已鉴定了2个重要的开花素基因Hd3a和RFT1,它们分别是短日照和长日照条件下促进抽穗的主效基因。在短日条件下,开花素Hd3a与14-3-3受体蛋白互作形成Hd3a/14-3-3复合物,然后与茎顶端分生组织中的OsFD1蛋白结合形成成花素激活复合物促进水稻成花转换。然而,水稻在长日照条件下成花素激活复合物形成的分子机制尚不清楚。
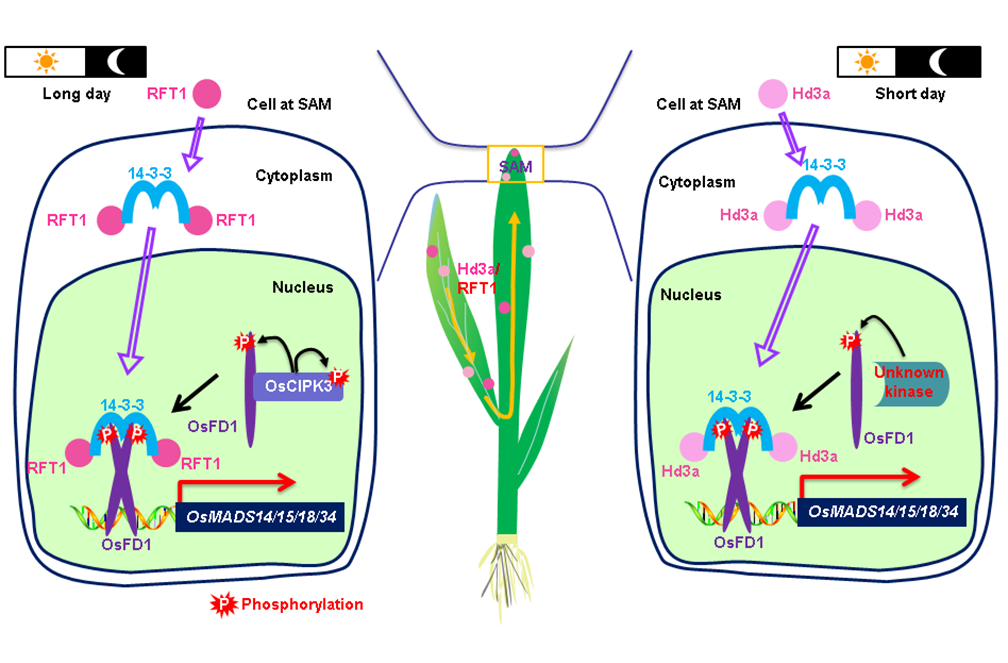
水稻成花素激活复合物形成的分子模型
研究人员通过酵母双杂交、GST pull-down、BIFC等实验,发现开花素RFT1通过14-3-3蛋白与磷酸化的成花启动基因OsFD1蛋白互作形成三元成花素激活复合物,从而行使在长日照条件下启动水稻成花转换的功能。研究发现,OsFD1中192位丝氨酸(S192)残基的磷酸化修饰对成花素激活复合物形成至关重要。进一步利用蛋白互作实验鉴定了一个负责对OsFD1磷酸化的蛋白激酶OsCIPK3。生化分析表明,OsCIPK3蛋白具有蛋白激酶活性,且证明OsCIPK3可以磷酸化修饰OsFD1蛋白的第192位丝氨酸位点。OsFD1的磷酸化修饰有利于成花素激活复合物进入细胞核,促进水稻由营养生长向生殖生长的转换。
遗传分析表明,OsCIPK3突变体表现为在长日照条件下晚抽穗,但短日照条件下抽穗期与野生型一致。OsCIPK3突变明显增加了穗的分枝数和小穗数,田间试验单株产量增加40%以上,表明OsCIPK3基因具有重要的育种利用价值。进一步通过生化实验分析表明,OsCIPK3主要负责在长日照条件下对OsFD1蛋白的磷酸化修饰,短日照条件下可能由其他的蛋白激酶参与OsFD1的磷酸化修饰,从而形成成花素激活复合物。
